Поиск по сайту
- Вы здесь:
-
Главная

-
История
-
Великие лекарства
-
book
-
Группы крови человека
- Система Gerbich
Наш блог
Это странная ситуация: вы соблюдали все меры предосторожности COVID-19 (вы почти все время дома), но, тем не менее, вы каким-то образом простудились. Вы можете задаться...
Как диетолог, я вижу, что многие причудливые диеты приходят в нашу жизнь и быстро исчезают из нее. Многие из них это скорее наказание, чем способ питаться правильно и влиять на...
Овес-это натуральное цельное зерно, богатое своего рода растворимой клетчаткой, которая может помочь вывести “плохой” низкий уровень холестерина ЛПНП из вашего организма....
Если вы принимаете витаминные и минеральные добавки в надежде укрепить свое здоровье, вы можете задаться вопросом: “Есть ли лучшее время дня для приема витаминов?”
Ты хочешь жить долго и счастливо. Возможно, ты мечтал об этом с детства. Хотя никакие реальные отношения не могут сравниться со сказочными фильмами, многие люди наслаждаются...
Приседания и выпады-типичные упражнения для укрепления мышц нижней части тела. Хотя они чрезвычайно распространены, они не могут быть безопасным вариантом для всех. Некоторые...

Ученые из Стэнфордского университета разработали программу предсказывающую смерть человека с высокой точностью.

Глава Минздрава РФ Вероника Скворцова опровергла сообщение о падении доходов медицинских работников в ближайшие годы. Она заявила об этом на встрече с журналистами ведущих...

Федеральная служба по надзору в сфере здравоохранения озвучила тревожную статистику. Она касаются увеличения риска острой кардиотоксичности и роста сопутствующих осложнений от...

Соответствующий законопроект внесен в палату на рассмотрение. Суть его заключается в нахождении одного из родителей в больничной палате бесплатно, в течении всего срока лечения...
- Категория: Система Gerbich
В систему Gerbich (Гербич) входят 8 антигенов, 3 из них Щ часто встречающиеся, 5 встречаются редко (табл. 20.1). Они располагаются в сиалопшкопротеинах мембраны эритроцитов, получивших название гликофорин С (GPC) и гликофорин D (GPD). Гликофорин D представляет собой укороченный вариант гликофорина С.
Синтез указанных двух гликофоринов контролируется одним геном - GYPC. В нем имеются два инициирующих участка, обеспечивающих синтез длинных и укороченных цепей, - GPC и GPD соответственно. Ген GYPC в составе четырех экзонов размещен на хромосоме 2 в позиции ql4—q21.
Часто встречающиеся антигены Gerbich (Ge2, Ge3 и Ge4) представлены соответствующими аминокислотными последовательностями. Редко встречающиеся антигены (Wb, Апа и Dha) являются результатом отдельных точковых мутаций в различных участках гена GYPC. Редкий антиген Lsa возникает вследствие дупликации экзона 3 указанного гена (см. табл. 20.1).
Таблица 20.1
Антигены Gerbich*
|
Обозначение |
Относительная частота, % |
Локализация на гликофорине |
|
|
традиционное |
ISBT |
||
|
Ge2 |
GE2 |
>99 |
N-терминальный участок GPD |
|
Ge3 |
GE3 |
>99 |
Аминокислоты в позициях 42-50 GPC и 21 - 29 GPD |
|
Ge4 |
GE4 |
>99 |
N-терминальный участок GPC |
|
Webb |
GE5 |
<1 |
Asn 8 Ser GPC |
|
Lsa |
GE6 |
<1 |
GPC и GPD, кодируемый GYPC с дупликацией экзона 3 |
|
Ana |
GE7 |
<1 |
Ala 2 Ser GPD |
|
Dha |
GE8 |
<1 |
Leu 14 Phe GPC |
|
GEIS |
GE9 |
<1 |
Thr 32Asn GPC и Thr 11 Asn GPD |
* Daniels [24], Issitt, Anstee [41], Reid и соавт. [97, 99].
Выделяют 3 редких фенотипа Gerbich, для которых характерно отсутствие одного антигена и более: Ge:-2,3,4 (фенотип Yussef, сокращенноYus), Ge:-2,-3,4 (фенотип Gerbich) и Ge:-2,-3,-4 (фенотип Leach). Указанные три фенотипа являются следствием трех вариантов делеций: экзона 2, экзона 3, экзонов 3 и 4 одновременно.
Лица, не содержащие антигенов Ge2, Ge3 и Ge4, встречаются, в основном, среди аборигенов Меланезии и Австралии. Редкие антигены обнаружены у представителей всех рас. Большинство антител системы Gerbich - IgM естественного происхождения, некоторые из них (aHTH-Ge2 и анти-веЗ) относятся к IgG. Трансфузионных осложнений и ГБН антитела Gerbich не вызывают.
- Категория: Система Gerbich
К гликофоринам относят гликопротеины с высоким содержанием сиаловых кислот. Они идентифицируются посредством электрофореза в додецилсульфат-ном полиакриламидном геле (SDS-PAGE). Большая часть гликофоринов мембраны эритроцитов (более 90 %) представлена вариантами А и В. Эти гликофорины несут антигены системы MNS.
Меньшую по величине долю (около 7 %) составляют два других высокогомологичных варианта гликофорина - С и D. Их представительство в мембране эритроцитов составляет 6 и 1 % соответственно (Furthmayr [33]). Гликофорины С и D несут антигены Gerbich.
Гликофорины А и В не связаны с гликофоринами С и D. Их синтез контролируют гены, расположенные на разных хромосомах.
Гликофорин С известен как протеин CD236C, р- и у-сиалогликопротеин, PAS-2'. Гликофорин D имеет синонимы: компонент D и компонент Е.
Мол. масса гликофоринов С и D 40 и 30 кДа соответственно (Reid [92], Colin, Le Van Kim [17], King [46]). Исследования с использованием Fab-фрагментов МКА к антигенам Gerbich показали, что один эритроцит содержит 143 тыс. молекул гликофорина С и 82 тыс. молекул гликофорина D (Smythe и соавт. [115]).
Расшифрована аминокислотная последовательность гликофоринов С и D (рис. 20.1) (Colin и соавт. [19], Dahr и соавт. [20]). аминокислотные последовательности трансмембранного домена, двумя линиями -участки связывания для протеинов 4.1R и р55 соответственно.

Аминокислотная последовательность гликофоринов С и D
Гликофорин С имеет три домена. Первый из них, экстрацеллюлярный N-терминальный гликозилированный (позиции 1-57) содержит N-связанный олигосахарид в положении Asn 8 и 12 участков О-гликозилирования. Второй домен - трансмембранный гидрофобный (позиции 58-81), третий домен -цитоплазматический С-терминальный (позиции 82-128) (Colin и соавт. [19], High, Tanner [38]). Цитоплазматический домен гликофорина С связан с ци-тоскелетоном. N-терминальный участок ассоциирован с трансмембранными гликофоринами А и В.
Аминокислотная последовательность гликофорина D определена частично, поскольку N-терминальный участок его блокирован (El-Maliki и соавт. [32]). Гликофорин D представляет собой укороченный вариант гликофорина С без N-терминальной последовательности из 21 аминокислоты. Он идентичен гликофорину С по аминокислотам в позициях 22-128.
Гликофорин D не имеет участков N-гликозилирования, поскольку не содержит N-терминального домена (Dahr и соавт. [21]). Антиген Ge3, представленный аминокислотной последовательностью в позициях 40-50 на гликофорине С, присутствует также на гликофорине D (High и соавт. [39]);
Антигенные детерминанты, распознаваемые ксеногенными моно- и поликлональными антителами, расположены на цитоплазматических доменах гликофоринов С и D.
- Категория: Система Gerbich
В 1960-х годах полагали, что существует один антиген Gerbich, выявляемый сывороткой миссис Gerbich и двумя другими однотипно реагирующими сыворотками. Этот антиген был описан Rosenfield и соавт. [106] и позднее получил обозначение Ge3. В 1961 г. Barnes и Lewis [6] нашли еще один антиген этой системы - Ge2, выявляемый сывороткой миссис Yussef. И наконец, в середине 80-х годов Anstee и соавт. [5], Daniels и соавт. [26], McShane и Chung [67] обнаружили антитела, идентифицирующие третий часто встречающийся антиген - Ge4.
Ge2, Ge3 и Ge4
Как установили Barnes и Lewis [6], эритроциты миссис Yussef, жительницы Кипра турецкого происхождения, агглютинировались сывороткой Gerbich, но не реагировали с сыворотками анти-Ge, полученными от других лиц Ge-. ыворотка миссис Yussef содержала антитела, реагировавшие со всем образцами эритроцитов за исключением собственных эритроцитов и эритроцитов трех ранее найденных Ge-отрицательных лиц. Адсорбция сыворотки Gerbich эритроцитами Yussef полностью устраняла ее активность. Таким образом, с помощью двух разнореагирующих сывороток (Gerbich и Yussef) были идентифицированы два антигена системы Gerbich.
Обследование жителей Папуа - Новой Гвинеи позволило получить новые данные об этой системе. Booth и соавт. [10, 11] нашли Gerbich-ассоциированные антитела у меланезийца Ge+, которые не реагировали с эритроцитами лиц, имевших фенотип Gerbich и Yus, а также с эритроцитами 15 % меланезийцев Ge+. Стало очевидным, что существует третья разновидность анти-Ое-антител, с помощью которой можно идентифицировать три Ge-отрицательных фенотипа, каждый из которых лишен одного из трех антигенов, составляющих систему Gerbich (табл. 20.3).
Таблица 20.3
Редкие фенотипы Gerbich
|
Носительница антител |
Фенотип |
Специфичность антител |
|
Mrs. Yussef |
Ge:-2,3,4 |
aHTH-Ge2 |
|
Mrs. Gerbich |
Ge:-2,-3,4 |
aHTH-Ge3 |
|
Mrs. Leach |
Ge:-2,-3,-4 |
aHTH-Ge4 |
Однако эритроциты меланезийца Ge:-1,2,3 и его анти-Gel-сыворотка вскоре стали недоступны. Соответственно, меланезийский вариант Gerbich-отрицательного фенотипа (Ge:-1,2,3) и анти-Gel-антитела не вошли в классификацию и больше не используются (Booth и соавт. [11,61], Daniels [24]).
Anstee и соавт. [5] и Daniels и соавт. [26] обнаружили, что некоторые исследованные ими моноклональные антитела обладали Gerbich-ассоциированной специфичностью. Эти антитела реагировали со всеми образцами эритроцитов Ge+ и Ge-, включая эритроциты Ge:-2,-3. В то же время были найдены образцы эритроцитов Ge:-2,-3, которые не реагировали с Gerbich-ассоциированными МКА (Anstee и соавт. [4, 5]). Этот Gerbich-отрицательный фенотип получил название Leach. Антитела, имевшиеся у миссис Leach, давали реакции, сходные с реакциями Gerbich-ассоциированных МКА. Антиген, открываемый ими, получил обозначение Ge4 (McShane, Chung [67]). Фенотип Leach обозначен в цифровой номенклатуре как Ge:-2,-3,-4. Все другие образцы эритроцитов содержат антиген Ge4 (см. табл. 20.3).
Посредством иммуноблоттинга с использованием нескольких образцов анти-Ое2-антител показано, что антиген Ge2 распологается на гликофорине D (рис 20.6). Гликофорин С не содержит этот антиген (Dahr и соавт. [22], Reid и соавт. [94]). Эпитопы Ge2 расположены на N-терминальном пептиде гликофорина D (аминокислоты в позициях 1-27) (Dahr и соавт. [22]). Обработка эритроцитов трипсином или папаином разрушает антиген Ge2, в то время как химотрипсин и проназа на него не действуют (Daniels [25]). Примерно половина образцов анти-Ое2-антител реагировала слабее с эритроцитами, обработанными сиалидазой (Daniels [25]).
Поскольку гликофорин D является укороченным аналогом гликофорина С анти-Ое2-антитела могут распознать аминокислотную последовательность, если она находится на N-терминальном участке гликофорина D, но не на гомологичном участке гликофорина С. Вместе с тем в образовании эпитопа Ge2 могут участвовать свободные аминогруппы гликофорина D. Некоторые образцы анти-Ое2-антител не реагируют с эритроцитами, обработанными уксусным ангидридом. Это дает основание полагать, что аминогруппы также вовлечены в формирование эпитопов, распознаваемых анти-Ое2-антителами. Анти-Ое2-антитела, по-видимому, являются гетерогенными и распознают эпитопы на разных участках гликофорина D, в том числе на N-терминальном участке (Daniels и соавт. [28]).
Подобно антигену Ge2, антиген Ge3 разрушается трипсином. К химотрипси-ну и проназе он устойчив. В отличие от Ge2 антиген Ge3 устойчив к действию папаина (Mohammed и соавт. [70]). Соответственно эритроциты, обработанные папаином, можно использовать для дифференциации антител aHTH-Ge2 от анти-Ge3 при отсутствии эритроцитов редкого фенотипа Ge:-2,3,4.
Методом иммуноблоттинга с МКА было показано, что эпитопы Ge3 локализуются на гликофоринах С и D (рис. 20.6) (Dahr и соавт. [22], Loirat и соавт. [56, 57], Reid и соавт. [94], Smythe и соавт. [115]). Аллоиммунные анти-веЗ-антитела удавалось элюировать с гликофоринов обоих типов (Reid и соавт. [94]).
Антиген Ge3 кодируется экзоном 3 GYPC. При делеций указанного участка антиген Ge3 отсутствует. Делеция экзона 2 не приводит к исчезновению экспрессии Ge3. Экзоны 2 и 3 имеют большое сходство и различаются лишь вставкой из 27 нуклеотидов, кодирующих аминокислоты в позициях 42-50 на гликофорине С и 21-29 - на гликофорине D, поэтому эпитопы Ge3 присутствуют именно в указанных областях соответствующих гликофоринов (Dahr и соавт. [22]).
Антиген Ge4 располагается в N-терминальной части гликофорина С (см. рис. 20.6). На гликофорине D он отсутствует (Anstee и соавт. [4, 5], Dahr и соавт. [21], Daniels и соавт. [29]).
Детальный анализ специфических МКА показал, что некоторым из них для связывания эпитопами Ge4 требовалась аминогруппа в позиции Metl на гликофорине С. В то же время другие МКА распознавали эпитопы среди первых 20 аминокислот гликофорина С и Metl в реакцию вовлечен не был (Anstee и соавт. [5], Dahr и соавт. [21]). Более значимым для связывания антител оказалось О-гликозилирование гликофорина С. Антиген Ge4 разрушается трипсином и папаином.
- Категория: Система Gerbich
Несмотря на высокую степень гомологии гликофоринов С и D, гомолог гена GYPC на уровне кДНК идентифицировать не удалось. Это свидетельствует о том, что самостоятельного гена, контролирующего синтез гликофорина D, не существует. Оба типа гликофоринов кодирует один и тот же ген GYPC (Le Van Kim и соавт. [52], Tanner и соавт. [120]). Как установили Tanner и соавт., различия гликофоринов С и D обусловлены мутацией иРНК гена GYPC, в результате чего трансляция начинается с двух разных точек, соответственно, синтезируются два гомологичных полипептида разной длины.

Неравновесный кроссинговер, приводящий к возникновению необычных вариантов гена GYPC
Структура гена GYPC
|
Экзон |
Позиции кодируемых аминокислот в гликофорине |
Локализация и специфичность антигена |
|
|
GPC |
GPD |
||
|
1 |
1-16 |
|
N-терминальный участок и часть экстра-целлюлярного домена GPC; N-гликан; Ge4 |
|
2 |
17-35 |
1—14 |
Met 22 на GPC, часть экстрацеллюлярной) домена GPC и GPD, включая его N-терминальный участок, Ge2 на GPD |
|
3 |
36-63 |
15-42 |
Часть экстрацеллюлярной) домена GPC и GPD, участок расщепления для трипсина на обоих гликофоринах, Ge3 |
|
4 |
64-128 |
43-107 |
Трансмембранный и цитоплазматические домены обоих гликофоринов |
Трансляция начинается с AUG-кодона (метионина), представленного в ДНК триплетом AEG. Вновь синтезированные полипептиды имеют метионин в N-области, хотя последний отщепляется от зрелого протеина. Последовательность РНК вблизи стартового кодона гликофорина С (CCAGGAAUGU) отдалена от стартовой последовательности (CC(A/G) CCAUGG). Последовательность (CCGGGGAUGG) кодона метионина в позиции 22 на гликофорине С находится ближе к стартовому участку (Tanner и соавт. [120]). Таким образом, первая стартовая точка (для метионина в позиции 1).
Второй участок, с которого может начаться трансляция, кодирует метионин в позиции 22. В этом случае синтезируется гликофорин D, который, как отмечалось выше, идентичен гликофорину С в позициях 22-128 (см. рис. 20.3). Возможность двух вариантов трансляции одного и того же гена подтверждена экспериментами с трансфек-цией кДНК GYPC в клетки COS-7. Указанные клетки продуцировали оба типа гликофоринов. Трансфекция этой линии кДНК с делециями ATG (кодона метионина) приводила к продукции только гликофорина D. В случае замены ATG на ACG в позиции 22 синтезировался гликофорин С. В случае замены обоих ко-донов ATG в позициях 1 и 22 гликофорины не синтезировались. Мутация в нуклеотиде 4 (ATG Т pi ATG G) повышала экспрессию гликофорина С в 2 раза по сравнению с выраженностью гликофорина D (Le Van Kim и соавт. [54]).
Ген GYPC представлен четырьмя экзонами общей протяженностью 13,5 кб (см. табл. 20.2, рис. 20.2) (Colin и соавт [18], High и соавт. [39]).
Экзоны 1-3 кодируют экстрацеллюлярные домены гликофоринов С и D, экзон 4 - трансмембранный и цитоплазматический домены. Экзоны 2 и 3 имеют
высокую степень сходства, что объясняется происхождением экзонов путем дупликации, хотя при этом экзон 3 содержит вставочную последовательность из 27 пар нуклеотидов, которой нет в экзоне 2. Указанная вставка кодирует аминокислоты в позициях 42-50 гликофорина С (Colin и соавт. [16]), Le Van Kim и соавт. [53]).
На рис. 20.4 и 20.5 представлена схема неравновесного кроссинговера и варианты гена GYPC, приводящие к возникновению редких антигенов и фенотипов Gerbich.
- Категория: Система Gerbich
Gerbich-отрицательные фенотипы у европеоидов крайне редки. При обследовании более 45 тыс. европейцев Gerbich-отрицательный фенотип выявлен лишь у одного (табл. 20.4). Существенно чаще люди, лишенные антигенов Gerbich, встречаются среди жителей Папуа - Новой Гвинеи.
Многие Gerbich-отрицательные лица выявлены в связи с присутствием в сыворотке их крови антител Gerbich (Anstee и соавт. [5], Barnes, Lewis [6], Daniels
и соавт. [29, 3], McLouglin, Rogers [66], Muller и соавт. [74], Nunn и соавт. [78], Okubo и соавт. [81], Peddle и соавт. [87], Reid и соавт. [91, 98], Rosenfield и соавт. [106], Rountree и соавт. [107], Sacks и соавт. [108]).
Фенотип Yus (Ge: -2,3,4) встречается реже, чем Ge:-2,-3, но такое соотношение может отражать лишь неодинаковую способность лиц с указанным фенотипом к антителообразованию (Daniels [25], Reid и соавт. [91]).
Лица Ge:-2,3,4 были обнаружены среди арабов, турок-киприотов и евреев, а также у негров.
Исследования посредством SDS-PAGE и иммуноблоттинга с МКА показано, что эритроциты Ge:-2,3,4 не содержат гликофорины С и D. Однако они несут гликофорин-С-подобную структуру с мол. массой 32,5-36,5 кДа, промежуточную между гликофоринами С и D (Anstee и соавт. [5], Dahr и соавт. [23], Reid и соавт. [94]). В эритроцитах гетерозигот GYPC'/GYPC. Yus гликофорины С и D присутствуют (Reid и соавт. [ 102]).
Анализ геномной и кодирующей ДНК показал, что лица Ge:-2,3,4 гомозиготны по гену GYP С. Yus, в котором экзон 2 подвергся делеций (Chang и соавт. [13], High и соавт. [39], Johnson, Daniels [44]). Продуктом указанного гена является гликофорин-С-подобный протеин без аминокислот в позициях 17-35, экспрессия антигенов Ge3 и Ge4 при этом сохранена. Вторая стартовая точка трансляции (Met 22) отсутствует, поэтому не происходит синтеза гликофорина D, несущего антиген Ge2.
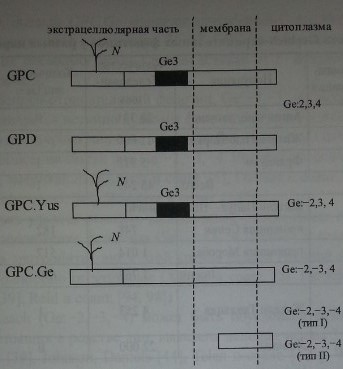
Локализация эпитопов Ge2-4 на гликофорине С и D
Частота Gerbich-отрицательных фенотипов у разных народов
|
Реагент, использованный при обследовании |
Популяция |
Количество обследованных |
Количество лиц Ge- |
Источник |
|
AHTH-Ge2 |
Англичане, датчане |
28 331 |
[66,90,106] |
|
|
Анти-веЗ |
Жители Нью-Йорка |
11000 |
[106] |
|
|
|
Французы |
5 912 |
1 |
[74,90] |
|
Всего: |
45 243 |
1 |
|
|
|
AHTH-Ge2 |
Жители Папуа - Новой Гвинеи: |
|||
|
|
провинция Сепик |
748 |
182 |
[П] |
|
|
провинция Моробе |
1014 |
517 |
|
|
|
Всего: |
1 762 |
699 |
|
|
Анти-Ge (точная специфичность неизвестна) |
Жители Таиланда |
4 253 |
1 |
[12] |
|
Анти-Ое2 |
Японцы |
22 ООО |
[81] |
|
Фенотип Ge:-2, 3, 4 может быть также результатом гетерозиготности по GYPC.Yus/GYPC.Ge (Loirat и соавт. [58], Moulds и соавт. [72]). У 5 из 10 лиц Ge:-2, 3 определялись оба типа гликофоринов - Yus и Ge (Moulds и соавт. [72]).
Фенотип Ge:-2,-3,4 является типичным Gerbich-отрицательньш. Он оказался полиморфным в некоторых районах Папуа - Новой Гвинеи (Booth, McLouglin и соавт. [11]). В других географических зонах земного шара фенотип Ge:-2,-3 встречается крайне редко, лишь в единичных случаях он был найден у европеоидов и негроидов, а также у монголоидов (японцев) и австралоидов (полинезийцев). Посемейные исследования показали, что гены, обусловливающие фенотип Ge:-2,-3, передаются по наследству (Nunn и соавт. [78], Okubo и соавт. [81], Reid и соавт. [100,102], Rosenfield и соавт. [106]).
Подобно эритроцитам Ge:-2,3,4, эритроциты Ge:-2,-3 не содержат гликофорины С и D, однако несут гликофорин-С-подобную структуру GPC.Ge с мол. массой 30,5-34,5 кДа, несколько меньшей по сравненю с мол. массой GPC.Yus. По электрофоретической подвижности вещество GPC.Ge занимает промежуточное положение между гликофоринами С и D (Anstee и соавт. [5], Dahr и соавт. [23], Reid и соавт. [94]). Гликофорин Gerbich-типа (GPC.Ge) содержит антиген Ge4, антиген Ge3 в нем отсутствует (Anstee и соавт. [5], Reid и соавт. [94]). В отличие от гликофоринов С, D и GPC.Yus, GPC.Ge устойчив к действию трипсина (Anstee и соавт. [5]). Моноклональные антитела к GPC.Ge (aHTH-Ge4) агглютинируют эритроциты Ge:-2,-3,4, обработанные трипсином. Они также агглютинируют эритроциты лиц, гетерозиготных по гену Ge~2,~3,4.
Фенотип Ge:-2,-3,4 возникает в результате делеций экзона 3 GPC (Colin и соавт. [18], Chang и соавт. [13], High и соавт. [39], Loirat и соавт. [58], Serjeantson и соавт. [ПО]). Аллель GPC.Ge кодирует гликофорин-С-подобную структуру, лишенную аминокислот в положениях 36-63. GPC.Ge несколько меньше гликофорина GPC.Yus, поскольку экзон 2 лишен вставки из 27 нуклеотидов, присутствующей в экзоне 3.
Еще один Gerbich-отрицательный фенотип, Ge:-2,-3,-4, получил обозначение Leach. Он является истинно нулевым фенотипом: эритроциты лиц с указанной редкой группой крови лишены всех антигенов системы Gerbich. В литературе имеются описания шести лиц Ge:-2,-3,-4. Все они оказались европеоидами (англичане, американцы) (Anstee и соавт. [5], Daniels и соавт. [30], McShane, Chung [67], Reid и соавт. [98], Rountree и соавт. [107]).
В двух семьях удалось показать наследование рецессивных генов, обусловливающих возникновение фенотипа Ge:-2,-3,-4 (Daniels и соавт. [30], Reid и соавт. [98]). На эритроцитах лиц с этой крайне редкой группой крови гликофорины С и D полностью отсутствуют (Anstee и соавт. [5], Daniels и соавт. [30], High и соавт. [39], Reid и соавт. [94, 98]).
Фенотип Leach (Ge:-2,-3,-4) может быть обусловлен разными причинами. У 5 не состоящих в родстве лиц, выявлена делеция экзона 3 и 4 гена GYPC (High и соавт. [39], Johnson, Daniels [44], Telen и соавт. [121], Winardi и соавт. [129]). В то же время из ретикулоцитов лиц Ge:-2,-3,-4 были выделены обычные транскрипты гена GYPC (Winardi и соавт. [129]).
У другого индивида Ge:-2,-3,-4 выделен полноценный ген GYPC, однако при секвенировании экзона 3 отмечена мутация, образующая стоп-кодон в позиции 56 (Telen и соавт. [121]). При этом не транслировалась большая часть экзона 3 и весь экзон 4. Тем не менее у обследуемых был выявлен небольшой фрагмент с мол. массой 12 кДа, напоминавший терминальную часть гликофоринов С и D (Pinder и соавт. [88]). В связи с этим высказано предположение, что трансляция дефектного гена GYPC может начинаться в других точках.
Имеется несколько сообщений о низкой экспрессии на эритроцитах Ge:-2,-3 антигенов Kell. Степень подавления у индивидов Ge:-2,-3 варьировала (Anstee и соавт. [5], Daniels и соавт. [25, 30], McShane, Chung [67], Nash и соавт. [77], Okubo и соавт. [81]). Снижение экспрессии отмечено в отношении часто встречающихся антигенов Kell, в также антигена KEL1. У одного индивида отмечена слабая экспрессия антигена KEL11 (Daniels [25]), выраженность других антигенов соответствовала норме. В 9 из 11 образцов эритроцитов Ge:-2,-3 отмечена слабая выраженность антигенов Kell. В то же время шесть образцов эритроцитов Ge:-2,3 имели нормальную экспрессию этих антигенов.
У 4 лиц Ge:-2,-3,-4 экспрессия антигенов Kell была слабой (Anstee и соавт. [5], Daniels и соавт. [30], McShane, Chung [67]).
Фенотипическая зависимость антигенов Kell от системы Gerbich реализуется, по-видимому, не на генетическом уровне, поскольку гены, контролирующие синтез вещества Gerbich и вещества Kell, расположены на разных хромосомах и непосредственное влияние их друг на друга исключено.


{kind=link}